My new paper in PeerJ features a new phylogenetic analysis of the Aetosauria (Archosauria: Pseudosuchia). I've added many new characters and feature all known valid aetosaur taxa. In the Supplemental Materials, each character is described and figured to clarify them for future use.

This paper also introduces a new aetosaur, Scutarx deltatylus, from the Chinle Formation of Arizona. To date I've collected four partial Scutarx skeletons, the first in 2002, from Petrified Forest National Park. The holotype specimen includes a partial skull, the first good aetosaur skull material collected from Arizona since the 1930s. In previous papers I have assigned all of these specimens to Calyptosuchus (Stagonolepis) wellesi; however, when I undertook a revision of that taxon I noted that my Petrified Forest specimens (all from the Sonsela Member of the Chinle Formation) all possessed raised triangular bosses on the posteromedial corner of the paramdian osteoderms. These are not present in the type specimen from Texas, or from the large amount of material from the Placerias Quarry of Arizona. Interestingly the boss-bearing and non-boss-bearing specimens were separated stratigraphically. Presently the specimens from the Blue Mesa Member of Arizona and the Tecovas Formation of Texas are those that lack the boss (Calyptosuchus wellesi), and those from the Sonsela Member, and middle Cooper Canyon Formation are those that possess the boss (Scutarx deltatylus). The skull and ilium of Scutarx are also autapomorphic as discussed in the paper.

Finally I adapted the method to compute Partioned Bremer Support to look at the possibility of character conflict between anatomical partitions in aetosaurs. Essentially does the armor have a different phylogenetic signal than the rest of the skeleton. Bremer Support is used in all phylogenetic analyses to show branch support; however, what is generally not realized is that the individual support values of characters from each anatomical partition combine to total this number. But if these character sets actually support a different tree than the one recovered in the total character analysis, they will reduce the Bremer Support value (negative Bremer Support). Some character sets are ambivalent and actually provide no support to the branches. This analytic tool allows you to look at your tree support node by node and see what character datasets support the topology. This technique has been used since the late 1990s to compare molecular vs. morphological character sets, but this is the first time it has been employed to look at anatomical partitions in a purely morphological study.
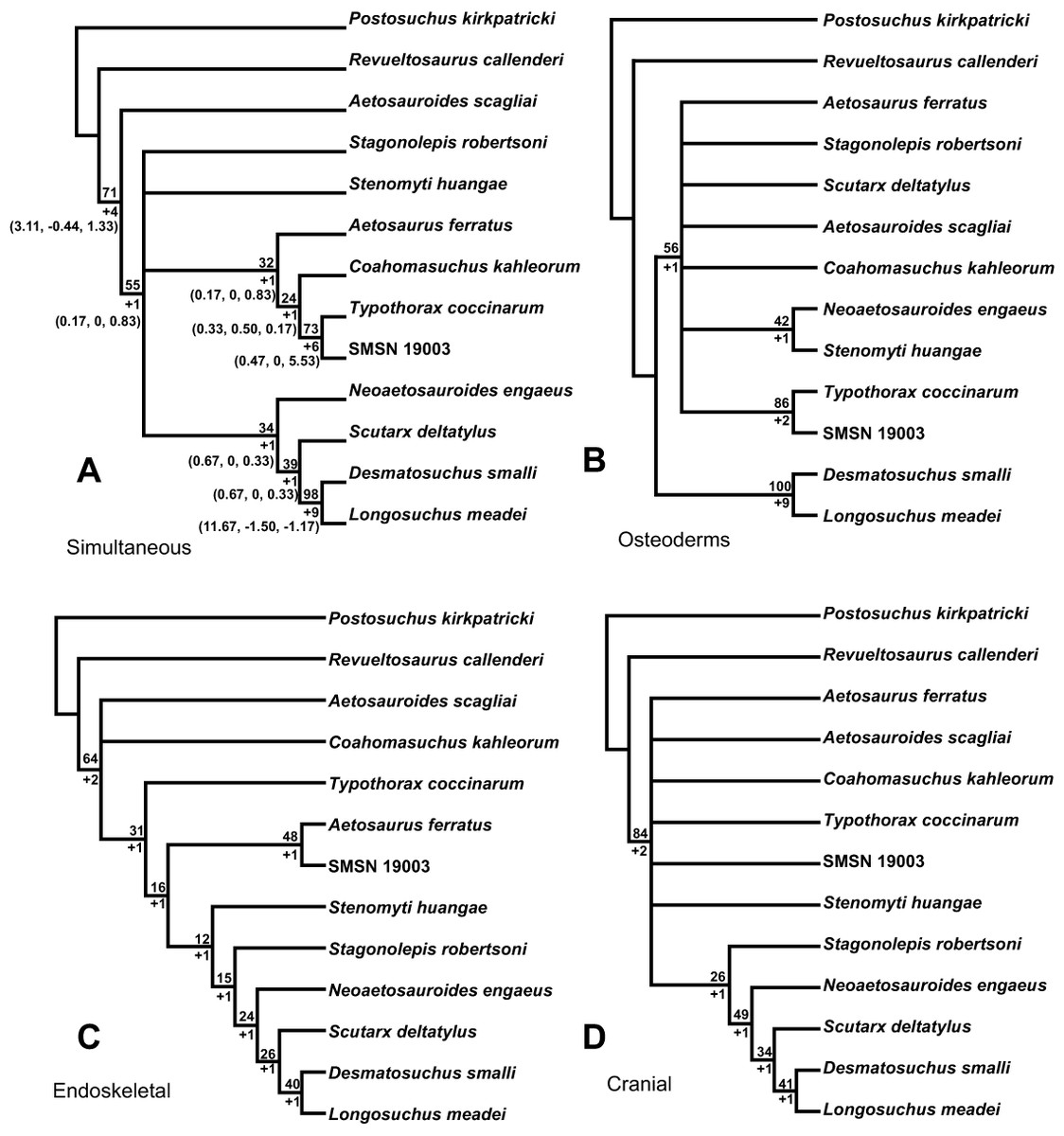
That said, I am a little disappointed with my final discussion section in the paper. While I am excited about the potential for Partitioned Bremer Support, I still am learning all that it demonstrates, thus my discussion is not as strong as I would have liked. I hope that others will find it of interest and utility and that we can see if if it works for other taxonomic groups.